
Published in Western Birds 52(1): An update on the status of the Anna’s Hummingbird in Idaho, as well as documentation of the first breeding record for the state
by Bryce W. Robinson
I’m privileged to see my name on two publications in Western Birds 52(1), alongside some great people from the Intermountain Bird Observatory. Both papers are focused on the changes to the status of Anna’s Hummingbird in Idaho, and feature some paintings and a photograph of mine.
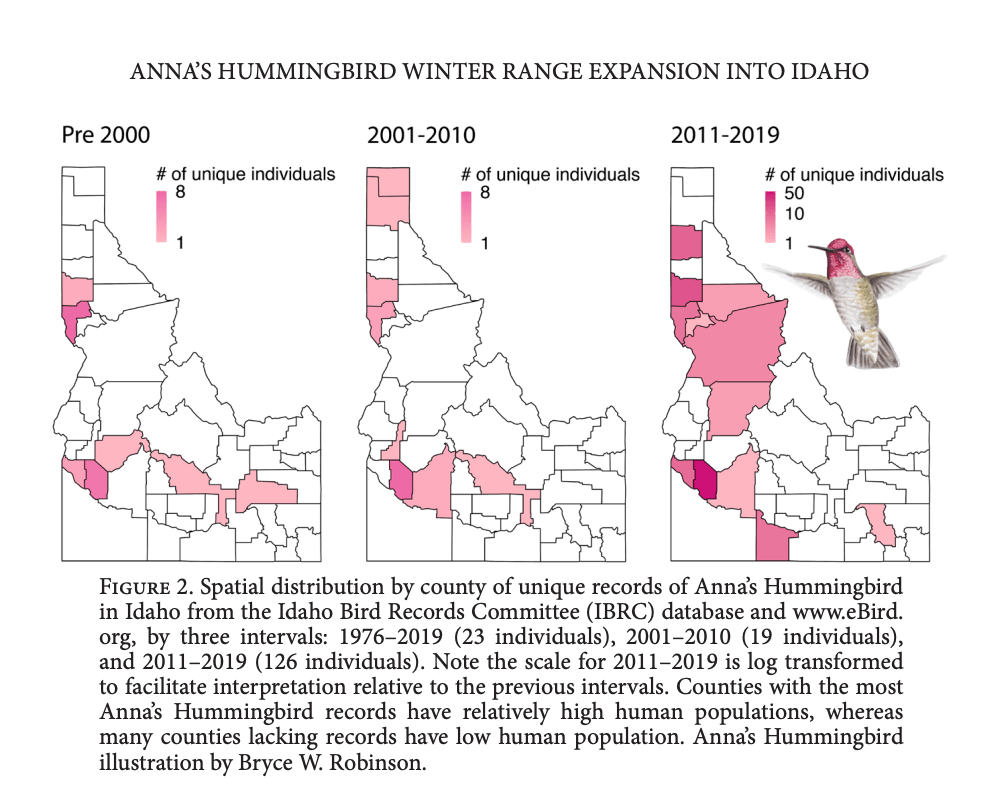
The main point from the first article led by Jessica Pollock – Number of over-wintering Anna’s Hummingbird in Idaho has substantially increased in the last ten years, and the species is now present year round in the state, perhaps as a resident.
The main point from the second article led by me – Documentation of the first successful breeding for the species in Idaho, as well as a discussion on why we can be reasonably sure these offspring are pure Anna’s and do not represent hybridization with a Black-chinned Hummingbird




